

Pigmentos y fotones: la ciencia detrás de los colores otoñales del bosque
A medida que los días se vuelven más fríos y aparecen las primeras escarchas, árboles y arbustos de hoja caduca comienzan el despliegue otoñal de tonos rojos, amarillos, púrpuras y marrones que caracterizan a los bosques templados de ambos hemisferios. Para comprender el proceso de diseño del fantástico espectáculo otoñal de los caducifolios es importante entender qué son y para qué sirven los pigmentos.


Artículo de Manuel Peinado Lorca (Catedrático de Universidad, Departamento de Ciencias de la Vida e Investigador del Instituto Franklin de Estudios Norteamericanos, Universidad de Alcalá) y Luis Monje (Biólogo, Profesor de fotografía científica, Universidad de Alcalá)
La magia química y lumínica de los pigmentos
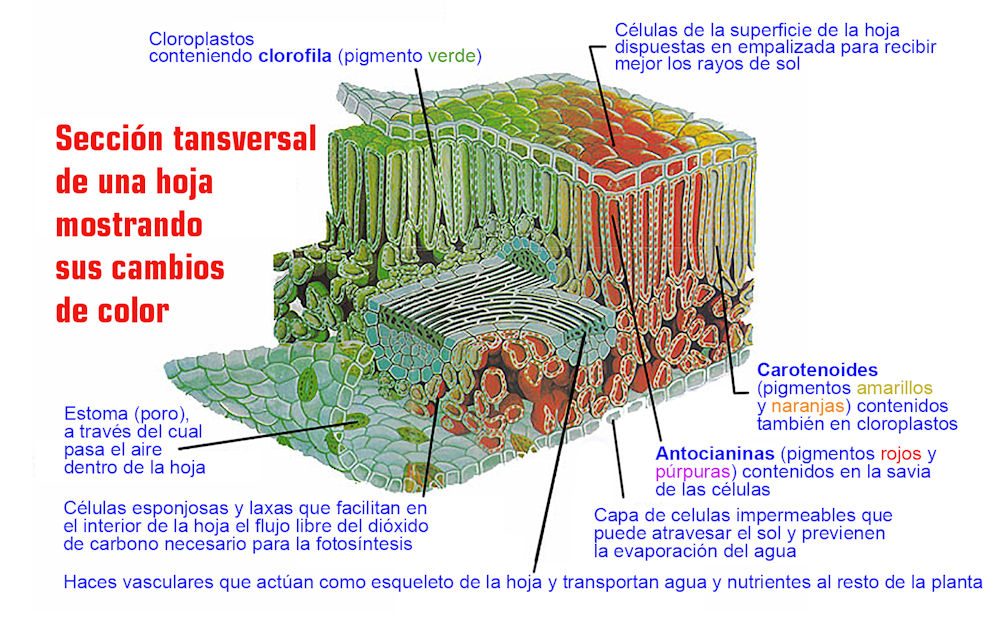
Las plantas son expertas en capturar la energía de la luz y utilizarla para crear azúcares mediante la fotosíntesis. Este proceso comienza con la absorción de luz mediante moléculas orgánicas especializadas, los pigmentos, que se encuentran en los cloroplastos y en las vacuolas celulares (Figura 1).
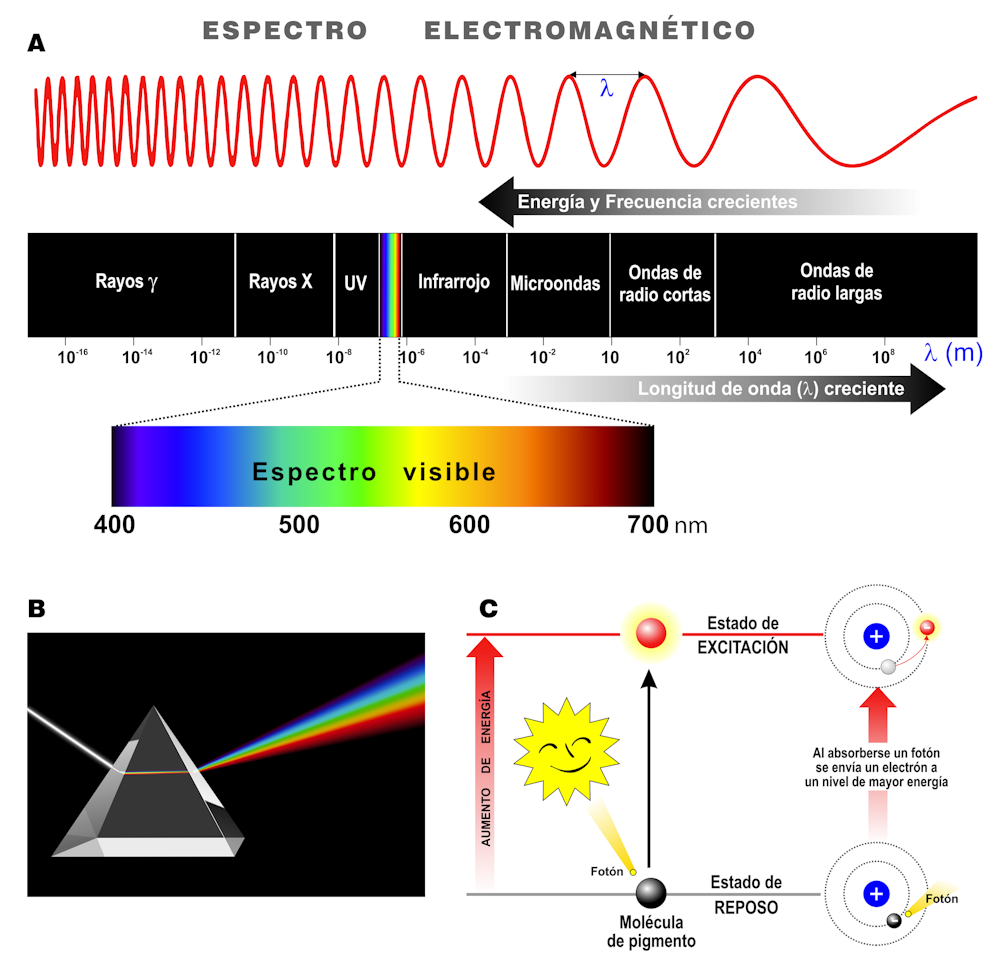
La luz es una forma de radiación electromagnética, un tipo de energía que viaja en ondas. En conjunto, todos los tipos de radiación conforman el espectro electromagnético, cuyas longitudes entre 400 y 700 nm constituyen la luz visible para el ojo humano (Figuras 2A y 2B). Cada partícula, llamada fotón, tiene una cantidad fija de energía que puede excitar un pigmento (Figura 2C).
Un pigmento excitado es inestable y tiene varias opciones disponibles para llegar a ser más estable. En las plantas, la energía de los fotones se usa para dividir moléculas de agua dentro de los cloroplastos. Además de oxígeno, en el proceso se liberan electrones e iones de hidrógeno. Estos electrones e iones se utilizan para generar energía en forma de adenosin trifosfato (ATP), que se usa en el ciclo de Calvin, cuyo objetivo es tomar CO₂ y utilizar la energía generada para transformar las moléculas de carbono en cadenas de moléculas orgánicas.
El color de un pigmento es el resultado de la longitud de onda reflejada. El color verde de las hojas se debe a la presencia de las clorofilas, unos pigmentos que se encuentran dentro de los cloroplastos, que absorben la mayoría de las radiaciones del espectro visible, y reflejan los verdes (Figura 3). Cuando abunda en las células, como sucede durante la temporada de crecimiento de primavera e inicios de verano, el color de la clorofila domina y enmascara los colores de cualquier otro pigmento que pudiera existir en la hoja.
La clorofila tiene una función vital: captura los rayos solares y utiliza la energía resultante en la fabricación de los alimentos de la planta: azúcares simples. Durante la temporada de crecimiento, la clorofila, que se deteriora con la exposición a la luz solar, se elabora, se degrada y se reemplaza constantemente en las hojas, fabricantes incansables de nueva clorofila durante la pujante estación de crecimiento.
El ciclo vital de los caducifolios
Como la mayoría de las plantas, los ritmos circadianos de los caducifolios son sensibles al fotoperiodo, es decir, a la duración del período diario de oscuridad. A finales del verano, los días comienzan a acortarse y las noches son más largas. Como la hora de inicio de todo el proceso depende de la duración de la noche, los colores otoñales aparecen aproximadamente a la misma hora cada año en un lugar determinado, sin que importe demasiado que las temperaturas sean más frías o más cálidas de lo normal.
Cuando las noches alcanzan un valor umbral y son lo suficientemente largas, las células que se encuentran en el punto de unión del peciolo de la hoja y el tallo que la sustenta se dividen rápidamente, pero no se expanden. Se forma así una capa de abscisión de callosa, es decir, una capa impermeable de células que poco a poco va obturando el floema y, con ello, bloqueando el transporte de savia de la hoja a la rama. También bloquea el xilema y, por tanto, el flujo de minerales desde las raíces hacia las hojas.
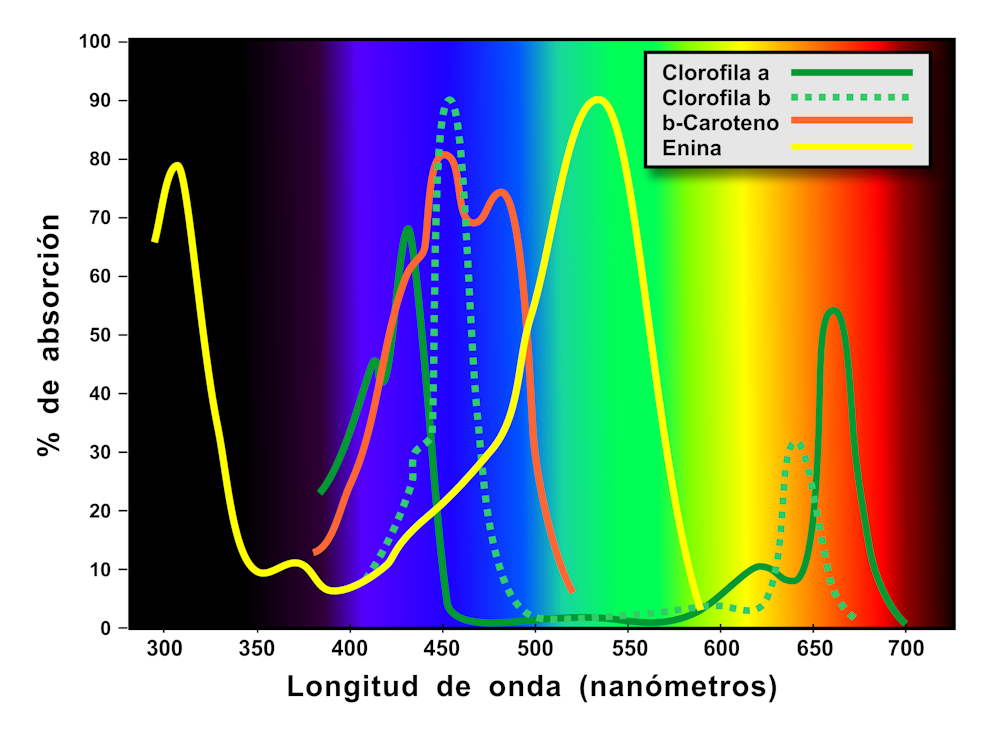
Conforme se desarrolla esta capa, la cantidad de clorofila en la hoja comienza a disminuir. A medida que las clorofilas se degradan, se revelan los pigmentos ocultos de unos derivados oxigenados de los carotenoides, las xantofilas amarillas, y los betacarotenos naranjas (Figura 3). Estos pigmentos están presentes durante todo el año en el interior celular, pero los pigmentos rojos, las antocianinas, se sintetizan de novo una vez que se ha degradado aproximadamente la mitad de la clorofila.
Aunque existan en las hojas durante todo el año, los colores amarillo-anaranjados de los carotenoides permanecen enmascarados por la clorofila verde. A medida que se acerca el otoño y el suministro total de clorofilas va disminuyendo gradualmente, el efecto de enmascaramiento se desvanece lentamente. Cuando eso sucede, comienzan a verse las coloraciones amarillas, pardas, naranjas y muchas tonalidades intermedias que proporcionan los carotenoides.
Los carotenoides son el pigmento dominante en la coloración de aproximadamente el 15-30 % de las especies de árboles. Sus amarillos y naranjas brillantes tiñen las hojas de nogales, fresnos, arces, álamos, abedules, cerezos, plátanos de paseo y alisos, entre otros muchos.
Los rojos, los púrpuras y sus combinaciones provienen de otro grupo de pigmentos celulares, las antocianinas. Hay dos diferencias importantes entre estos pigmentos y las clorofilas y los carotenoides. La primera es que no se encuentran en los cloroplastos. Son pigmentos solubles en agua que se almacenan en las vacuolas. La segunda es que, a diferencia de los carotenoides, no existen en la hoja durante la temporada de crecimiento, sino que se producen activamente hacia el final del verano.

Cuando se acerca el otoño, las antocianinas se elaboran a partir de la savia acumulada en las células. Su síntesis de novo depende de la descomposición de los azúcares en presencia de luz intensa a medida que se reduce el nivel de fosfato en la hoja. Durante la temporada de crecimiento de verano, el nivel de fosfato es alto, porque juega un papel vital en la descomposición de los azúcares fabricados por la clorofila. En otoño, el fosfato, junto con otros nutrientes, se moviliza desde la hoja al tallo de la planta. Cuando eso sucede, el proceso de descomposición del azúcar cambia, lo que lleva a la producción de pigmentos de antocianina. Cuanto más brillante sea la luz durante este período, mayor será la producción de antocianinas y más luminoso será el color resultante.
En las regiones de clima templado, las antocianinas están presentes en aproximadamente una de cada diez especies de árboles aunque en algunos lugares hasta el 70 % de las especies de árboles pueden producir el pigmento. En los bosques otoñales tiñen a arces, robles, cornejos, cerezos y ciruelos. Estos mismos pigmentos a menudo se combinan con los colores de los carotenoides para crear los naranjas más intensos, los rojos vivos y los bronceados típicos de muchas especies de madera dura.
A medida que avanza el otoño, las células de la capa de abscisión se encallecen más y más. Como la clorofila, los otros pigmentos se degradan con la luz o cuando se congelan. Los únicos pigmentos que quedan son los taninos acumulados en las paredes celulares, que son marrones. Las conexiones entre las células se debilitan y las hojas acaban por caer.
Ha llegado el invierno y con él cae el maravilloso telón multicolor de la temporada otoñal que, en esencia, como toda la vida que nos rodea, está literalmente construida desde cero por la partícula que encumbró a Einstein: el fotón. Bastante increíble, ¿no está de acuerdo?




